
Cell:化学信号的“换班”,炎症如何改变免疫系统的导航机制
来源:生物探索 2024-12-24 14:38
该研究以极高的科学价值和应用潜力,推动了免疫学领域的进一步发展。
炎症是人体免疫系统应对感染或损伤的重要反应过程,而淋巴结作为免疫防线中的“前哨站”,在炎症中扮演着关键角色。当身体感染病毒或细菌时,局部淋巴结常会肿大,这是大量淋巴细胞迅速聚集的结果。这一现象在日常生活中并不少见,但背后的科学机制却异常复杂。淋巴细胞如何精准进入发炎的淋巴结,并有效启动免疫应答,一直是免疫学领域的重要研究课题。
在健康状态下,淋巴细胞依赖于一种名为CCR7(CC-chemokine receptor-7)的受体与其配体CCL21之间的相互作用,通过高内皮小静脉(high endothelial venules, HEVs)进入淋巴结。然而,当炎症发生时,这一机制却发生了显著变化。
12月20日Cell的研究报道“Inflammation switches the chemoattractant requirements for naive lymphocyte entry into lymph nodes”揭示,在炎症环境中,CCL21的表达显著下调,而另一种化学信号分子,由胆固醇氧化生成的氧化甾醇(oxysterol)及其受体EBI2(GPR183),开始主导淋巴细胞的迁移过程。这种从“化学信号A”到“化学信号B”的转变,为淋巴细胞的募集提供了一条全新的路径。
更令人惊讶的是,研究人员发现,这一过程背后隐藏着多种细胞和分子的精密协作。炎症状态下,高内皮小静脉和淋巴结中的成纤维细胞协同工作,分泌了氧化甾醇,为淋巴细胞提供了关键的指引。同时,一些免疫细胞如朗格汉斯细胞(Langerhans cells)也参与了这一复杂网络,进一步促进了淋巴细胞的招募。
这一发现不仅为我们理解炎症状态下免疫系统的运行机制提供了重要线索,也为开发针对炎症相关疾病的新疗法打开了大门。通过调控这些信号分子和通路,我们或许能够有效增强抗感染能力,或控制过度活跃的免疫反应,例如在自身免疫疾病或肿瘤环境中。

炎症与免疫:淋巴结的“警报中心”
在人体复杂的免疫防御网络中,淋巴结如同一座高效的“指挥中心”,时刻守护着我们的健康。人体内约有500个淋巴结,分布在关键的解剖位置,形成了一个严密的免疫监视网。它们通过接收来自外周组织的淋巴液,将潜在威胁如病原体或肿瘤细胞快速传递给免疫细胞,触发全面的免疫反应。
淋巴结的功能如同警报器,能够精准识别“敌人”的信号。当外界病原体入侵或组织受到损伤时,炎症信号会通过局部淋巴管被传递至淋巴结。在这里,成千上万的淋巴细胞,包括B细胞和T细胞,迅速聚集并被激活,为清除病原体或修复受损组织做准备。研究表明,在健康状态下,淋巴细胞通过高内皮小静脉(high endothelial venules, HEVs)进入淋巴结,这一过程依赖于CCR7受体与其配体CCL21的精确互动。
然而,当炎症来袭,情况变得更加复杂。炎症诱发的化学信号会改变淋巴结内部环境,增强其招募淋巴细胞的能力。具体而言,炎症状态下的淋巴结会表达更多氧化甾醇(oxysterol)等信号分子,以吸引淋巴细胞的到来。这一策略确保了更多种类的免疫细胞能够参与到免疫反应中,提高了识别和清除病原体的效率。该研究揭示,在病毒感染的早期阶段,炎症淋巴结中的淋巴细胞迁移速率显著提升,表明淋巴结动态调节其免疫功能以适应外界环境的变化。
更重要的是,淋巴结的这种“警报中心”角色并不仅仅限于抗感染。它们还在调节自身免疫反应、控制肿瘤进展和应对疫苗接种中发挥重要作用。理解淋巴结在炎症中的关键作用,不仅帮助我们揭开免疫系统的运行机制,更为改善临床治疗提供了新思路。例如,如何通过调控淋巴细胞迁移来增强疫苗效能,或减少自身免疫疾病中的过度免疫反应,都是值得深入探索的问题。
淋巴细胞如何找到路?化学趋化因子揭示的秘密
健康状态下,淋巴细胞在血液和淋巴结之间的往返穿梭,是免疫系统高效运转的重要保障。然而,这些“小卫士”如何精准找到淋巴结中的位置并完成任务?答案隐藏在一套精妙的化学导航系统中,其中的关键角色是CCR7受体和CCL21趋化因子。
CCR7(CC-chemokine receptor 7)是一种表达在B细胞和T细胞表面的化学受体,而CCL21则是其高亲和力的配体,主要由高内皮小静脉(high endothelial venules, HEVs)和淋巴结中的成纤维细胞分泌。在正常情况下,CCL21通过形成明确的化学梯度,为循环中的淋巴细胞指引方向。这一“化学地图”不仅确保淋巴细胞能够从血液进入淋巴结,还能够引导它们在淋巴结内的精确定位,使其抵达T细胞区域或B细胞滤泡,准备迎接来自外界的抗原挑战。
研究显示,这种导航系统的核心是CCR7与CCL21的动态互动。当CCR7感受到CCL21的信号后,会启动细胞内部的信号级联反应,触发淋巴细胞的趋化性运动,使其朝着化学梯度的高浓度区域迁移。该研究进一步证实,在CCR7缺失的小鼠中,淋巴细胞无法有效进入淋巴结,导致免疫反应显著减弱。这表明,CCR7和CCL21系统是淋巴细胞迁移过程中不可或缺的基础模块。
更有趣的是,这一系统并非孤立存在。其他趋化因子如CXCL12和CXCL13,也参与了某些特殊免疫细胞的导航工作,与CCR7-CCL21形成互补网络。这种多维度的趋化因子系统,确保了不同种类的免疫细胞能够在需要时迅速聚集到特定位置。
淋巴细胞的迁移不仅是一个生物化学过程,更是免疫系统协调运作的典范。正是因为这套化学信号导航系统的存在,免疫系统才能在平衡与效率间找到最佳点。而当炎症发生时,这一平衡又会如何改变?
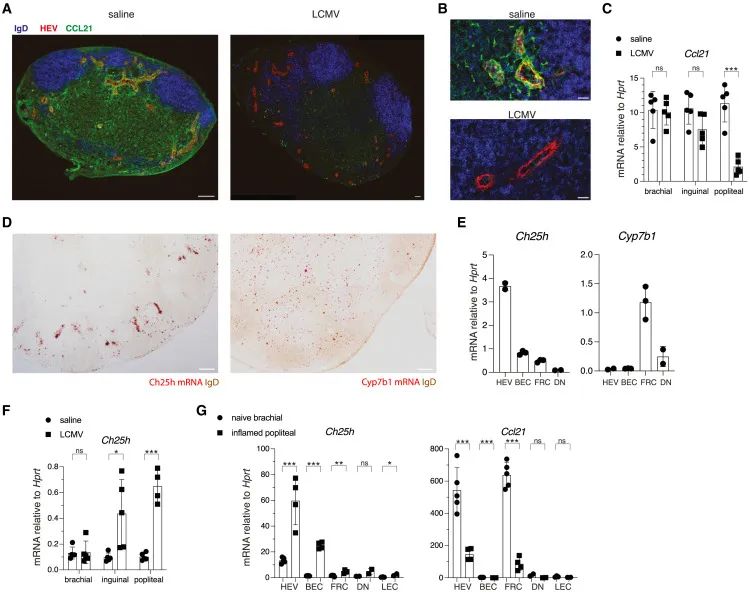
感染期间淋巴结基质细胞下调CCL21并上调Ch25h(Credit: Cell)
(A) IFM图像:感染对CCL21表达的影响
显示了未感染的小鼠(左)和感染24小时后小鼠足底淋巴细胞性脉络脑膜炎病毒(LCMV)引流淋巴结(pLN,右)的免疫荧光染色结果。标记的区域包括高内皮小静脉(PNAd+ HEVs)、IgD阳性B细胞区域和CCL21表达区域。感染后,CCL21的信号显著减少,特别是在HEVs和T细胞区的表达几乎消失。
(B) HEVs的放大图
进一步放大的高内皮小静脉图像揭示,在未感染的淋巴结(顶部),CCL21表达强烈,而在感染后(底部),CCL21的表达几乎完全下调。
(C) RT-qPCR分析CCL21表达
实时定量PCR数据显示,在感染的引流淋巴结(如popliteal LN)中,CCL21的mRNA水平显著降低,而在未受感染的对照淋巴结(如brachial LN)中未观察到类似变化。这表明CCL21下调与感染直接相关。
(D) Ch25h和Cyp7b1的RNA-scope图像
RNA-scope实验显示,在感染引流的pLN中,Ch25h的表达显著上调,尤其是在高内皮小静脉和T细胞区域的外周。同时,Cyp7b1更多地局限于T细胞区。这种分布表明,Ch25h和Cyp7b1在淋巴结的不同区域协作发挥功能。
(E) 不同细胞中的Ch25h和Cyp7b1表达
通过对不同淋巴结基质细胞(如高内皮小静脉细胞(HEVs)、血管内皮细胞(BECs)、成纤维细胞(FRCs)和双阴性基质细胞(DN cells))的RT-qPCR分析,发现Ch25h在HEVs中高表达,而Cyp7b1在成纤维细胞中显著表达,表明这两种细胞类型在氧化甾醇信号生成中具有分工。
(F) Ch25h的整体上调
RT-qPCR数据显示,感染引流的淋巴结中Ch25h的表达显著高于未感染淋巴结,进一步验证了感染刺激Ch25h上调的事实。
(G) 基质细胞中的Ch25h和CCL21表达比较
在感染引流的pLN和未感染的brachial LN中分离的基质细胞(如成纤维细胞)中,RT-qPCR数据显示感染时Ch25h显著上调,而CCL21则显著下调,验证了CCL21与Ch25h的表达模式具有相反趋势。
炎症来袭:化学信号的“换班”机制
当炎症来袭,淋巴结不再是平静的免疫枢纽,而是变成了战时的“指挥中心”。在这一过程中,淋巴细胞迁移的化学信号系统经历了显著变化,标志着一场“化学信号的换班”。研究发现,健康状态下主导淋巴细胞进入淋巴结的CCL21趋化因子,在炎症状态下显著下调,而一种新的信号轴,氧化甾醇(oxysterol)与其受体EBI2(GPR183),迅速兴起,成为主导力量。
该研究揭示,在病毒感染后的早期阶段,CCL21的表达在淋巴结的高内皮小静脉(HEVs)和T细胞区域成纤维细胞中急剧减少,降幅高达6至8倍。而与此相对的是,HEVs中一种关键酶胆固醇-25-羟化酶(cholesterol-25-hydroxylase, Ch25h)的表达显著上调。这种酶催化生成了氧化甾醇7α,25-二羟基胆固醇(7α,25-HC),而该分子正是EBI2受体的高效配体。研究表明,这一转变能显著增强淋巴细胞迁移至发炎淋巴结的效率。
这一机制的背后,体现了多种细胞类型的精密协作。高内皮小静脉产生的7α,25-HC通过跨细胞代谢途径,与淋巴结成纤维细胞表达的Cyp7b1酶协同工作,形成了淋巴细胞导航的全新化学梯度。此外,朗格汉斯细胞(Langerhans cells)等免疫细胞也在炎症条件下贡献了部分氧化甾醇的生成。
为什么需要这一“信号换班”?答案是效率。CCL21在正常状态下足以维持基础的免疫监控,但在炎症环境中,大量淋巴细胞需要快速到位,以提高抗原识别的概率。这时,Ch25h-EBI2轴的兴起,为淋巴细胞提供了更快速、更精确的迁移路径。
“团队合作”:多细胞协作生成淋巴细胞导航信号
在炎症状态下,淋巴细胞迁移的效率依赖于一种精密的“团队合作”。高内皮小静脉(HEVs)、成纤维细胞和朗格汉斯细胞(Langerhans cells)等多种细胞协同作用,共同生成并维持了氧化甾醇信号轴(Ch25h-EBI2),为淋巴细胞的导航提供精准指引。
研究表明,HEVs在炎症环境中是氧化甾醇合成的核心动力。它们表达高水平的胆固醇-25-羟化酶(Ch25h),催化胆固醇生成25-羟基胆固醇(25-HC)。这一分子随后扩散至淋巴结内的其他细胞中,例如成纤维细胞,在这些细胞中,酶Cyp7b1进一步将25-HC转化为7α,25-二羟基胆固醇(7α,25-HC),最终为淋巴细胞提供导航信号。这种跨细胞的代谢协作,确保了炎症状态下氧化甾醇的高效生成和分布,形成了一个明确的化学梯度。
成纤维细胞作为淋巴结的结构性基质细胞,在这一过程中扮演了关键的辅助角色。它们不仅是Cyp7b1的主要表达来源,还通过维持淋巴结的组织完整性,确保了导航信号的有效传递。此外,研究显示成纤维细胞的这一功能在炎症环境中更为显著,与HEVs形成了紧密的功能耦合。
与此同时,朗格汉斯细胞等免疫细胞也加入了这一复杂网络。作为一种特殊的抗原提呈细胞,朗格汉斯细胞在炎症状态下显著上调Ch25h的表达,成为淋巴结中氧化甾醇的补充来源。实验数据还表明,这些细胞主要位于淋巴结的T细胞区域,与淋巴细胞接触频繁,从而强化了导航信号的局部效应。
这种“团队合作”机制,不仅揭示了炎症状态下多种细胞的协调能力,也凸显了氧化甾醇信号的动态特性。通过这一合作网络,淋巴细胞能够高效迁移到需要它们的地方,为免疫反应提供关键支持。这种机制的发现为治疗炎症相关疾病带来了新希望,比如通过靶向调控这些细胞的协作,可以更加精准地调节免疫反应。
不只是淋巴结:炎症对其他组织的影响
氧化甾醇信号并非淋巴结的“专属工具”。近年来的研究表明,这一信号轴在肿瘤和其他炎症相关组织中同样发挥了重要作用,为理解免疫系统的整体调控提供了新的视角。
在肿瘤环境中,炎症反应的特征之一是肿瘤周围的淋巴结(tumor-draining lymph nodes, tdLNs)的显著改变。研究发现,tdLNs中CCL21的表达下降,而氧化甾醇信号的成分,如Ch25h和EBI2,显著上调。这种变化有助于增强淋巴细胞的募集,使免疫细胞更高效地聚集在肿瘤周围并可能形成抗肿瘤的“第三方淋巴组织”(tertiary lymphoid structures, TLS)。这些组织被认为可以作为抗肿瘤免疫应答的关键调控点。该研究进一步显示,EBI2缺失的小鼠在肿瘤相关淋巴结和肿瘤本体中表现出显著的淋巴细胞迁移障碍,这表明氧化甾醇信号对肿瘤免疫具有不可替代的作用。
炎症不仅改变了淋巴结的内部环境,还对其他外周组织产生深远影响。在慢性炎症性疾病如特应性皮炎(atopic dermatitis)的患者皮肤中,氧化甾醇信号同样被激活。研究发现,皮肤内的微血管内皮细胞表达Ch25h,并生成氧化甾醇,为EBI2阳性的免疫细胞提供迁移信号。这种现象暗示,氧化甾醇信号可能是多种炎症性皮肤病的重要参与者,并为治疗这些疾病提供了新靶点。
不仅如此,该研究还提示,炎症引起的氧化甾醇信号轴变化可能广泛存在于外周组织,包括呼吸道和胃肠道的炎症环境。例如,在病毒感染后的肺部炎症中,氧化甾醇信号可以促进淋巴细胞的招募,从而提升局部的免疫防御能力。这种跨组织的调控机制表明,氧化甾醇信号具有全身性影响力,不仅限于特定的免疫组织。
因此,氧化甾醇信号为我们提供了一个理解炎症如何系统性调控免疫反应的窗口。这一发现不仅深化了对肿瘤免疫学的认知,还为开发治疗慢性炎症和肿瘤的新策略提供了重要启示。
双重保障:CCL19如何接替CCL21维持淋巴细胞迁移
在炎症中,免疫系统需要以最快的速度调集兵力,而这背后的关键在于化学信号的无缝衔接。当主导淋巴细胞迁移的CCL21在炎症中显著下调时,另一种化学趋化因子,CCL19,则迅速“接棒”,与氧化甾醇信号(Ch25h-EBI2轴)形成双重保障,确保淋巴细胞能够精准、高效地迁移至发炎的淋巴结。
该研究发现,在炎症状态下,尽管CCL21的表达显著降低,但CCL19的表达却保持稳定,甚至在某些条件下上调。作为CCR7的另一种配体,CCL19在淋巴结的T细胞区域成纤维细胞中高度表达。不同于CCL21主要集中于高内皮小静脉(HEVs),CCL19更广泛分布于淋巴结内部结构,这种空间分布的差异使得CCL19能够在炎症期间补充和扩展CCR7信号。研究表明,在CCL19缺失的小鼠中,炎症淋巴结的淋巴细胞迁移效率明显下降,特别是对于EBI2缺失的细胞来说,这一缺陷更为显著,凸显了CCL19在炎症条件下的非冗余作用。
更有趣的是,CCL19与氧化甾醇信号之间存在显著的协同效应。CCL19通过与CCR7结合,为淋巴细胞提供额外的化学导航信息,而氧化甾醇信号则进一步增强了这种迁移能力。研究显示,当同时阻断CCL19和氧化甾醇信号时,淋巴细胞的迁移几乎完全停止,表明两者共同构成了淋巴细胞进入发炎淋巴结的关键路径。此外,炎症状态下的HEVs表面对CCL19的结合能力增强,使其能够进一步覆盖CCL21下调后留下的信号“空白”。
这种双重保障机制的存在,为我们提供了免疫调控的另一层视角。CCL19不仅在维持炎症期间的淋巴细胞迁移中起到核心作用,还与氧化甾醇信号密切合作,共同为免疫细胞的精确定位提供动力。这一发现不仅深化了我们对化学趋化因子功能的理解,也为未来通过靶向调控CCR7或EBI2通路治疗炎症相关疾病提供了更多可能性。
未解之谜:是什么控制了CCL21的“沉默”?
CCL21是维持淋巴细胞迁移的重要信号分子,但在炎症状态下,它却经历了显著的表达下调。这一现象背后的机制仍是一个未解之谜。理解这种“沉默”现象,不仅对免疫调控具有重要意义,也为开发针对炎症的治疗策略提供了科学依据。
研究发现,CCL21在淋巴结中高内皮小静脉(HEVs)和成纤维细胞中的表达在炎症早期就急剧下降。这种下调可能与多种炎症信号通路有关。该研究表明,干扰素(interferons)在这一过程中发挥了关键作用,特别是I型干扰素(IFN-α/β)。在实验中,缺失干扰素受体的小鼠表现出显著减弱的CCL21下调,提示干扰素信号可能通过调控CCL21的转录或翻译过程发挥作用。然而,γ型干扰素(IFN-γ)在CCL21的调控中似乎作用有限,这表明不同干扰素信号可能具有特异性功能。
除了干扰素信号,炎症环境中的其他因素如细胞因子和代谢压力也可能参与其中。研究显示,炎症激活的核因子κB(NF-κB)信号可能抑制CCL21的基因表达。此外,淋巴结内的机械应力,如炎症期间淋巴液流量的变化,也被认为会影响CCL21的分泌水平。值得注意的是,CCL21的基因座在炎症状态下并未表现出显著的染色质封闭,这表明其下调可能更多地与转录后调控或蛋白降解有关,而非染色质结构的改变。
CCL21的“沉默”似乎有其独特的生物学意义。在炎症状态下,CCL21的下调为氧化甾醇信号(Ch25h-EBI2)和CCL19提供了更大的调控空间。这种动态切换不仅优化了淋巴细胞的迁移效率,也可能是为了避免信号的冗余或紊乱,从而使免疫系统更高效地应对外界威胁。
这一领域的研究仍有许多未解之谜。例如,是否存在特定的分子“开关”直接控制CCL21的表达?未来的研究或许可以探索更细致的分子机制,例如RNA结合蛋白或非编码RNA在CCL21调控中的角色,这将进一步揭示炎症期间免疫调控的复杂性。
重塑对免疫系统运行的理解
这项研究的发现以全新视角揭示了淋巴细胞迁移的动态调控机制,对免疫学领域具有里程碑式的意义。从健康状态下的CCR7-CCL21信号主导,到炎症状态下Ch25h-EBI2和CCL19的接替,这一系列研究重塑了我们对免疫系统运行的理解。它不仅深化了对淋巴结在免疫应答中角色的认知,还揭示了不同信号分子在不同条件下的功能转换,为探索炎症和免疫相关疾病的新疗法提供了重要依据。
研究中的核心成果在于揭示了免疫系统的高度灵活性和适应性。在健康状态下,淋巴细胞依靠CCR7-CCL21信号完成日常巡逻。然而,当炎症发生,CCL21表达下调,氧化甾醇信号和CCL19迅速兴起并接管淋巴细胞的导航工作。这种动态调控机制确保了免疫细胞在需要时能高效地聚集到发炎部位。研究不仅扩展了我们对趋化因子功能的理解,还强调了多细胞协作和跨细胞代谢的重要性。
此外,该研究为临床实践提供了新启示。通过调控氧化甾醇信号轴和CCL19,未来可以更精准地控制炎症反应,提高抗感染能力,或在自身免疫和肿瘤免疫治疗中降低病理性炎症和增强免疫应答。这一发现不仅为基础研究开辟了新领域,也为个性化医疗和免疫调节技术提供了新思路。
然而,许多谜团仍未解开。例如,CCL21在炎症中的下调具体由哪些信号调控?淋巴结内不同细胞间的协作是否存在其他未被发现的关键节点?以及在慢性炎症或肿瘤环境中,这些信号分子的动态变化是否不同?未来的研究可以结合高分辨成像和单细胞组学技术,深入探讨免疫细胞迁移的微观机制,同时挖掘更多潜在的免疫调控靶点。
总的来说,该研究以极高的科学价值和应用潜力,推动了免疫学领域的进一步发展。它不仅回答了关于淋巴细胞迁移的关键问题,还为探索免疫系统的复杂运行方式提供了全新的框架。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
